Introduction
The purpose of this symposium is to bring together the results of work done by
CSULB biogeographers on California sage scrub in Southern California. The
team has included three summers of field and lab work done by the NSF-funded
Geoscience Diversity Enhancement Program (or GDEP), led by Paul Laris and
myself, several master's theses, a biogeography graduate seminar, and several
sections of courses in biogeography, landscape restoration, and field methods.
My remarks will provide an overview and context.
[ Viewgraph
2 ] California sage scrub is a scrub vegetation dominated by smaller
shrubs and subshrubs that form a more open canopy than does chaparral. Its
adaptations to summer drought include a greater emphasis on pubescent leaves
and shallower root networks than typical in chaparral, and several species
have developed facultative summer deciduousness. [ Viewgraph 3 ]
The result is a visually distinct vegetation that in summer is predominantly
grey to tan. [ Viewgraph 4 ]
The bulk of it is found near the coast in Southern California, where fog drip
can be an important moisture source during the summer. [ Viewgraph 5 ]
A significant amount of it is also found inland away from the coast in such
locations as the San Fernando Valley, San Gabriel Valley, and western San
Bernardino and Riverside counties. It is commonly called coastal sage scrub
along the coast and interior sage scrub inland. We'll usually refer to the
whole complex as California sage scrub or CSS.
[ Viewgraph
6 ] CSS has markedly declined in Southern California in the face of
grazing and associated clearing, plowing, construction, residential
landscaping practices, changes in fire regimes, and air pollution. Dwindling
to an estimated 10-15% of its previous range, the loss of CSS has threatened a
number of endangered animal species with loss of habitat. [ Viewgraph 7
], [ Viewgraph
8 ], [ Viewgraph 9
], [ Viewgraph
10 ]. Among these are the California gnatcatcher (Polioptila
californica), Palos Verdes blue butterfly (Glaucopsyche lygdamus
palosverdesensis), the coastal cactus wren (Campylorhynchus
brunneicapillus), and several others. The belated recognition of the
vital habitat functions played by CSS has led to attempts to conserve and
restore it and to understand the forces converting it to other land cover
types.
[ Viewgraph
11 ] A common type conversion is from CSS to exotic-dominated annual
grassland, which is often strikingly able to maintain its dominance long past
the disturbance that originally led to its replacement of CSS. Several
hypotheses have been put forward to explain the persistence of the grassland
conversion.
CSS Suppression
[
Viewgraph
12 ] Fire regimes are altered by the new light fuel that can grow rapidly
in winter and spring, move quickly to reproduction, and then die off by
summer, encouraging the rapid spread of fire. More frequent fire favors
grasses, which have already completed their life cycles when summer and fall
fires strike and consume their dead biomass. More frequent fires can exceed
the ability of CSS to stump sprout and maintain seed banks dense enough to
regenerate the CSS species mix.
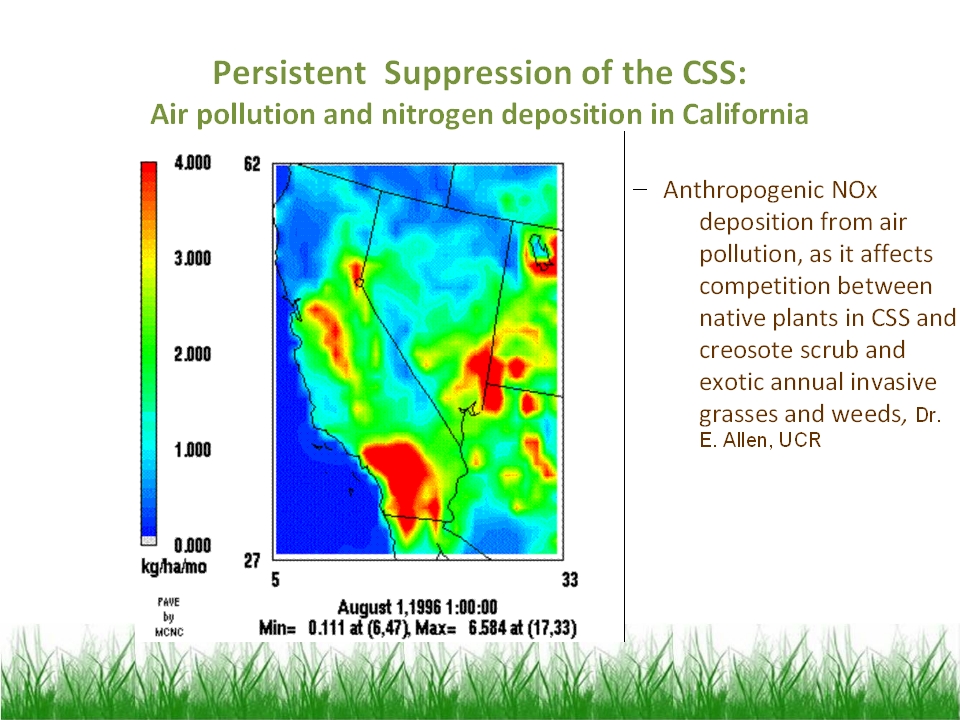
[ Viewgraph
13 ] Nitrogen fertilization has shown an intriguing connection to CSS
suppression and grassland persistence. Annual grasses and exotic forbs seem
better able to utilize nitrogen than native shrubs and forbs in CSS, and
native gardening nurseries strongly recommend not using nitrogen fertilizer.
Nitrogen fertilization occurs due to the deposition of nitrous oxides in air
pollution, and research on interior sage scrub has shown marked spatial
correlation between CSS degradation and grassland type conversion in San
Bernardino and Riverside counties. Our own work on Palos Verdes and the Santa
Monicas has not shown any such effect, perhaps because of the cleaner air by
the coast.
[ Viewgraph
14 ] Allelopathy has been posited to explain the persistence of
Brassica nigra and Hirschfeldia incana mustards in annual grassland and the
inability of CSS to recolonize mustard-infested grasslands. Our own work
evaluating the effect of mulched dead mustard stalks on the germination and
growth of California poppies proved inconclusive.
[ Viewgraph
15 ] Grazing has often been cited in the decline of CSS and the
persistence of grasslands. CSS was actively cleared to create rangeland and
pastures, and the combination of soil compaction, accelerated erosion,
uprooting by livestock, and active planting of forage species prevented
recolonization by CSS. The question was whether the damage would lead to
permanent type-conversion. Hearteningly, several studies, including those
done by the CSULB group, show that CSS can reclaim territory once grazing
ends.
[ Viewgraph
16 ] Less hearteningly are findings about the permanence of mechanical
disturbance of the soil, as by plowing. Our own work indicates that, if CSS
is cleared and the soil plowed even once, grasses and associated exotic forbs,
notably black mustard and fennel, will not relinquish a site to CSS. Paul
Laris found a site high on a steep slope near Del Cerro Park in Palos Verdes
that had been plowed once around 1928 and, though never used again, the field
is still discernible by the black mustard dominating it.
[ Viewgraph
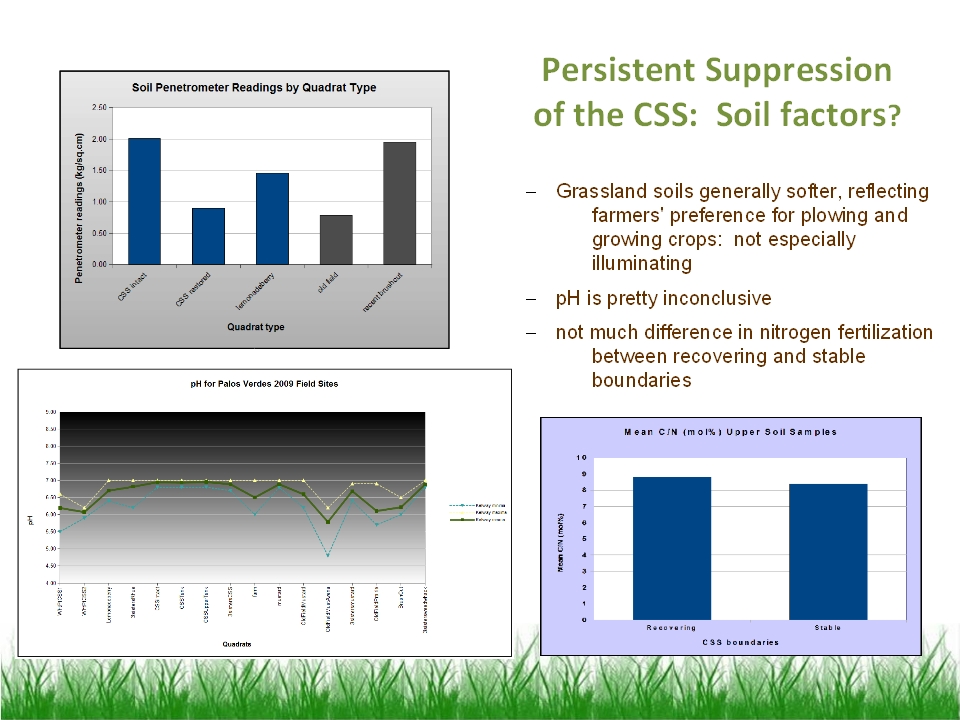
17 ] Soil factors may have something to do with it, but there's no
unambiguous signal. Soils occupied by grasslands tend to be softer and have a
higher clay and silt content than contemporary CSS soils. Whether this has
anything to do with CSS preference for edaphic conditions is moot: It is more
likely that farmers selected the softer and more fertile soils for their
fields and avoided rockier and more gravelly soils and outcrops to avoid
damaging their equipment. pH does not seem to vary much between CSS and
grassland, and nitrogen fertilization does not distinguish recovering from
stable boundaries.
[ Viewgraph
18 ] It is possible that the extreme persistence of grassland in once
plowed soils may reflect the disruption of subsurface symbioses between native
plant roots and various mycorrhizal fungus species. Weedy exotics and many
grasses get along fine without mycorrhizal infections and, so, may establish
and maintain dominance over a plowed site. Many native shrubs and forbs
strike up symbioses with particular mycorrhizal species and may be unable to
compete with exotics without their symbiotic partners. The mechanical
disturbance of plowing may destroy these subsurface relationships. One of the
papers in this session gives a progress report on evaluating this idea.
CSS Restoration
[
Viewgraph
19 ] Concerted efforts to restore CSS have often been disappointing, as
grasses and associated exotic forbs reclaim restoration sites. Conservancies
struggle with trying to find the right way to re-establish CSS, trying
different soil treatments, different palettes of plant species, various
irrigation systems, and a lot of weeding. There have been successes, such as
the Trump Golf Course, which is de facto an intensively gardened landscaping
project, way past the resources of most volunteer organizations. There have
been a lot of failed projects or projects that are only partially successful,
including one in the Tijuana watershed documented in a thesis by Samantha
Antcliffe. Active restoration can be a frustrating slog.
That said, there are a number of places where CSS has been able to encroach
back onto grasslands on its own. CSULB Geography became involved in this
through Scott Eckardt's thesis, which used air photos and remote sensing to
track CSS decline in Calabasas. He found patches of CSS that were actually
expanding in the face of overall decline. Kyra Engelberg worked in the
Serrano and La Jolla valleys in the westernmost Santa Monica Mountains,
identifying sites through air photos and satellite imagery where CSS and grass
had maintained an unchanging boundary for decades and others where CSS was
pushing into grassland. The GDEP team and subsequent classes have gone out
there transecting these border types at sites she selected, in order to
characterize their flora and succession processes and the relevance of slope,
soil factors, underlying geology, and subsurface communities. You'll be
hearing progress reports on much of this today.
Applications for CSS Active and Passive Restoration
[
Viewgraph
20 ] Understanding the reasons for CSS self-restoration is critical for
the success of landscape restoration programs. If we can identify conditions
correlated with CSS self-restoration, conservancies can identify sites in
their holdings or potential acquisitions where passive restoration might occur
with little demand on their scarce labor and material supplies.
There may be significant differences in the CSS species mix found behind a
stagnant boundary and behind an expanding boundary. Also, we have been
censusing CSS species found out in front of the expanding boundaries, CSS
species that can tolerate the conditions in the grasslands and perhaps serve
as nurses fostering other CSS species. It is possible that active restoration
efforts could be designed to mimic a succession process modeled on the
expanding CSS boundaries, planting only vanguard species at first, later
adding species found disproportionately behind expanding CSS boundaries, and
then adding the rest of the local CSS species palette.
A cautionary note emerges from the findings about the permanent type
conversion created by plowing and other forms of mechanical disturbance and
soil overturn. Some active restoration sites are prepared first by soil
scraping, which removes topsoil and weed seed banks, presumably to cut down on
exotic weed recruitment back to the site to give native plantings a fighting
chance. To the extent that this mechanically disturbs soil, it may lead to
enduringly poor outcomes. Soil scraping and plowing may have this unintended
consequence by disrupting mycorrhizal communities below ground.
If mycorrhizæ prove to be suppressed in grassland and ubiquitous in CSS,
or, alternatively, if mycorrhiza are disrupted in once plowed or scraped
sites, active restoration needs to inoculate new sites with appropriate
species of symbiotic fungus. Mycorrhizæ: are available for purchase
from suppliers Back East, but there is some evidence from a variety of
ecosystems that there are particular mutualisms involved and a generic
inoculum may not have the intended effect.
Rest of the Symposium
Kyra
Engelberg and Paul Laris will summarize transects taken in Serrano and La
Jolla valleys and present results about slope, soil texture, and species mixes
in the stable and shifting boundary zones (.ppt).
Paul
Nesbit and Scott Winslow will summarize a project they did to analyze the
association between geological substrate and stable CSS-grassland boundary
zones (.ppt).
Mystyn
Mills and Paola Ducoing Chaho will present work she's done on the presence
of mycorrhiza on rootballs and soil samples taken in La Jolla Valley (.pptx).
Sean
Brennan, Chelsea Vaughan, and Carin Tabag will summarize work they did in
the westernmost La Jolla Valley to assess the rôle of herbivory on the
expansion of CSS (.pptx).
Sean
Brennan will also present work he's done evaluating the rôle of
Baccharis pilularis as a vanguard species and CSS facilitator (.pptx).
Jade
Dean will outline the work she did on the differences in species mixes
with distances from the CSS-grassland boundary in both expanding and stable
boundaries (.pdf).
Paul
Laris will wrap up, summarizing the overall findings from the symposium
papers and from another paper given a week ago at another conference and
drawing out conclusions for land managers and restoration ecologists (.ppt).
Note: The other paper discussed by Laris was presented at the CSULB College
of Liberal Arts Student Research Showcase by Abel
Santana, Lourdes Johanna Avelar-Portillo, and Joseph Diminutto and
evaluated gravitational bias in CSS self-restoration (.odp).
Selected References
Allen, Edith B.; Padgett, Pamela E.; Bytnerowicz, Andrzej; and Minnich,
Richard. 1998. Nitrogen deposition effects on coastal sage vegetation of
Southern California.
USDA Forest Service General Technical Report
PSW-GTR-166: 131-139. Available at
http://gis.fs.fed.us/psw/publications/documents/psw_gtr166/psw_gtr166_002_allen.pdf.
.
Allen, Edith B.; Eliason, Scott A.; Marquez, Viviane J.; Schultz, Gillian P.;
Storms, Nancy K.; Stylinski, Cathlyn Davis; Zink, Thomas A.; and Allen,
Michael F. 2000. What are the limits to restoration of coastal sage scrub in
Southern California? In 2nd Interface between Ecology and Land Development
in California, ed. Jon E. Keeley, Melanie Baer-Keeley, and C.J.
Fotheringham. U.S. Geological Survey Open-File Report 00-62: 253-262.
Article available at http://www.faculty.ucr.edu/~eallen/allenetal_2000.html
(full anthology available as a 118 meg PDF at http://pubs.usgs.gov/of/2000/of00-062/of00-062.pdf).
Bell, David T., and Muller, Cornelius H. 1973. Dominance of California annual
grasslands by Brassica nigra. American Midland Naturalist 90, 2: 277-
299. JSTOR URL: http://www.jstor.org/stable/2424453.
Cox, Robert D., and Allen, Edith B. 2008. Stability of exotic annual grasses
following restoration efforts in southern California coastal sage scrub.
Journal of Applied Ecology 45, 2: 495-504.
doi: 10.1111/j.1365-2664.2007.01437.x.
Fleming, Genie M.; Diffendorfer, James E.; Zedler, Paul H. 2009. The
relative importance of disturbance and exotic-plant abundance in California
coastal sage scrub. Ecological Applications 19, 8: 2210-2227.
Hobbs, E.R. 1986. Characterizing the boundary between California annual
grassland and coastal sage scrub with differential profiles. Vegetatio
65: 115-126.
Keeley, Jon E.; Fotheringham, C.J.: and Baer-Keeley, Melanie. 2005.
Determinants of postfire recovery and succession in Mediterranean-climate
shrublands of California. Ecological Applications 15, 5: 1515-1534.
Stinson, Kristina A.; Campbell, Stuart A.; Powell, Jeff R.; Wolfe, Benjamin
E.; Callaway, Ragan R.; Thelen, Giles C.; Hallett, Steven G.; Prati, Daniel;
and Klironomos, John N. 2006. Invasive Plant suppresses the growth of native
tree seedlings by disrupting belowground mutualisms. PLOS Biology 4, 5:
e140. doi: 10.1371/journal.pbio.0040140. Available at http://www.plosbiology.org/article/info:doi/10.1371/journal.pbio.0040140.
Talluto, Matt V.; Suding, Katherine Nash; and Bowler, Peter A. 2006. Factors
affecting understory establishment in coastal sage restoration.
Madroño 53, 1: 55-59.
Vogelsang, Keith M., and Bever, James D. 2009. Mycorrhizal densities decline
in association with nonnative plants and contribute to plant invasion.
Ecology 90, 2: 399-407.
Yoshida, Lidia C., and Allen, Edith B. 2001. Response to ammonium and
nitrate by a mycorrhizal annual invasive grass and native shrub in Southern
California. American Journal of Botany 88, 8: 1430-1436.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}