COGNITIVE MAPS IN RATS AND MEN[1]
Edward C. Tolman (1948)
First published in The Psychological Review, 55(4), 189-208.
I shall devote the body of this paper to a description of experiments with
rats. But I shall also attempt in a few words at the close to indicate the
significance of these findings on rats for the clinical behavior of men. Most of
the rat investigations, which I shall report, were carried out in the Berkeley
laboratory. But I shall also include, occasionally, accounts of the behavior of
non-Berkeley rats who obviously have misspent their lives in out-of-State
laboratories. Furthermore, in reporting our Berkeley experiments I shall have to
omit a very great many. The ones I shall talk about were carried out by
graduate students (or underpaid research assistants) who, supposedly, got some
of their ideas from me. And a few, though a very few, were even carried out by
me myself.
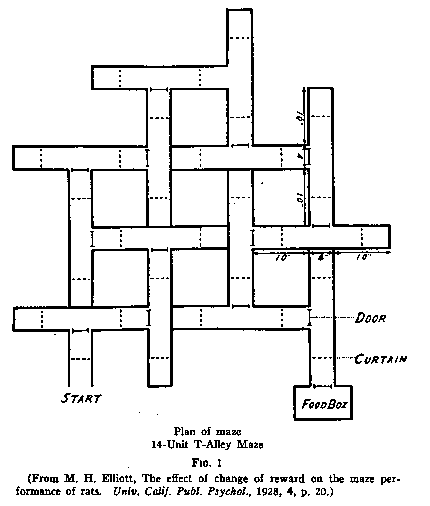
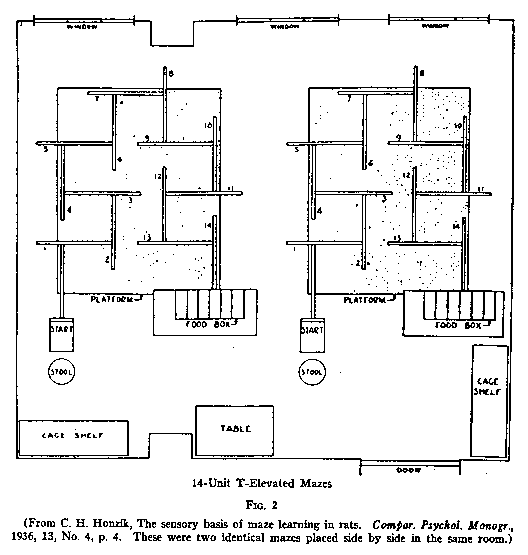
Let me begin by presenting diagrams for a couple of typical mazes, an alley maze
and an elevated maze. In the typical experiment a hungry rat is put at the
entrance of the maze (alley or elevated), and wanders about through the various
true path segments and blind alleys until he finally comes to the food box and
eats. This is repeated (again in the typical experiment) one trial every 24
hours and the animal tends to make fewer and fewer errors (that is, blind-alley
entrances) and to take less and less time between start and goal-box until
finally he is entering no blinds at all and running in a very few seconds from
start to goal. The results are usually presented in the form of average curves
of blind-entrances, or of seconds from start to finish, for groups of rats.
All students agree as to the facts. They disagree, however, on theory and
explanation.
(1) First, there is a school of animal psychologists which believes that the
maze behavior of rats is a matter of mere simple stimulus-response connections.
Learning, according to them, consists in the strengthening of some of these
connections and in the weakening of others. According to this
'stimulus-response' school the rat in progressing down the maze is helplessly
responding to a succession of external stimuli-sights, sounds, smells,
pressures, etc. impinging upon his external sense organs-plus internal stimuli
coming from the viscera and from the skeletal muscles. These external and
internal stimuli call out the walkings, runnings, turnings, retracings,
smellings, rearings, and the like which appear. The rat's central [p.190]
nervous system, according to this view, may be likened to a complicated
telephone switchboard.
There are the incoming calls from sense-organs and there are the outgoing
messages to muscles. Before the learning of a specific maze, the connecting
switches (synapses according to the physiologist) are closed in one set of ways
and produce the primarily exploratory responses which appear in the early
trials. Learning, according to this view, consists in the respective
strengthening and weakening of various of these connections; those connections
which result in the animal's going down the true path become relatively more
open to the passage of nervous impulses, whereas those which lead him into the
blinds become relatively less open.
It must be noted in addition, however, that this stimulus-response school
divides further into two subgroups.
(a) There is a subgroup which holds that the mere mechanics involved in the
running of a maze is such that the crucial stimuli from the maze get presented
simultaneously with the correct responses more frequently than they do with any
of the incorrect responses. Hence, just on a basis of this greater frequency,
the neural connections between the crucial stimuli and the correct responses
will tend, it is said, to [p.191] get strengthened at the expense of the
incorrect connections.
(b) There is a second subgroup in this stimulus-response school which holds
that the reason the appropriate connections get strengthened relatively to the
inappropriate ones is, rather, the fact that the responses resulting from the
correct connections are followed more closely in time by need-reductions. Thus a
hungry rat in a maze tends to get to food and have his hunger reduced sooner as
a result of the true path responses than as a result of the blind alley
responses. And such immediately following need-reductions or, to use another
term, such 'positive reinforcements' tend somehow, it is said, to strengthen the
connections which have most closely preceded them. Thus it is as if-although
this is certainly not the way this subgroup would themselves state it-the
satisfaction-receiving part of the rat telephoned back to Central and said to
the girl: "Hold that connection; it was good; and see to it that you
blankety-blank well use it again the next time these same stimuli come in."
These theorists also assume (at least some of them do some of the [p.192] time)
that, if bad results-'annoyances,' 'negative reinforcements'-follow, then this
same satisfaction-and annoyance-receiving part of the rat will telephone back
and say, "Break that connection and don't you dare use it next time
either."
So much for a brief summary of the two subvarieties of the
'stimulus-response,' or telephone switchboard school.
(2) Let us turn now to the second main school. This group (and I belong to
them) may be called the field theorists. We believe that in the course of
learning something like a field map of the environment gets established in the
rat's brain. We agree with the other school that the rat in running a maze is
exposed to stimuli and is finally led as a result of these stimuli to the
responses which actually occur. We feel, however, that the intervening brain
processes are more complicated, more patterned and often, pragmatically
speaking, more autonomous than do the stimulus-response psychologists. Although
we admit that the rat is bombarded by stimuli, we hold that his nervous system
is surprisingly selective as to which of these stimuli it will let in at any
given time.
Secondly, we assert that the central office itself is far more like a map
control room than it is like an old-fashioned telephone exchange. The stimuli,
which are allowed in, are not connected by just simple one-to-one switches to
the outgoing responses. Rather, the incoming impulses are usually worked over
and elaborated in the central control room into a tentative, cognitive-like map
of the environment. And it is this tentative map, indicating routes and paths
and environmental relationships, which finally determines what responses, if
any, the animal will finally release. [p.193]
Finally, I, personally, would hold further that it is also important to
discover in how far these maps are relatively narrow and strip-like or
relatively broad and comprehensive. Both strip-maps and comprehensive-maps may
be either correct or incorrect in the sense that they may (or may not), when
acted upon, lead successfully to the animal's goal. The differences between such
strip maps and such comprehensive maps will appear only when the rat is later
presented with some change within the given environment. Then, the narrower and
more strip-like the original map, the less will it carry over successfully to
the new problem; whereas, the wider and the more comprehensive it was, the more
adequately it will serve in the new set-up. In a strip-map the given position of
the animal is connected by only a relatively simple and single path to the
position of the goal. In a comprehensive-map a wider arc of the environment is
represented, so that, if the starting position of the animal be changed or
variations in the specific routes be introduced, this wider map will allow the
animal still to behave relatively correctly and to choose the appropriate new
route.
But let us turn, now, to the actual experiments. The ones, out of many, which I
have selected to report are simply ones which seem especially important in
reinforcing the theoretical position I have been presenting. This position, I
repeat, contains two assumptions: First, that learning consists not in
stimulus-response connections but in the building up in the nervous system of
sets which function like cognitive maps, and second, that such cognitive maps
may be usefully characterized as varying from a narrow strip variety to a
broader comprehensive variety.
The experiments fall under five heads: (1) "latent learning," (2)
"vicarious trail and error" or "VTE," (3) "searching
for the stimulus," (4) "hypotheses" and (5) "spatial
orientation."[p.194]
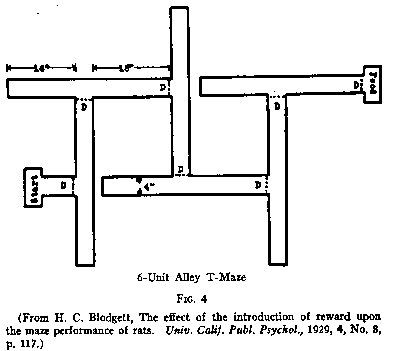
(1) "Latent Learning" Experiments. The first of the latent
learning experiments was performed at Berkeley by Blodgett. It was published in
1929. Blodgett not only performed the experiments, he also originated the
concept. He ran three groups of rats through a six-unit alley maze, shown in
Fig. 4. He had a control group and two experimental groups. The error curves for
these groups appear in Fig. 5. The solid line shows the error curve for Group I,
the control group. These animals were run in orthodox fashion. That is, they
were run one trial a day and found food in the goal-box at the end of each
trial. Groups II and III were the experimental groups.
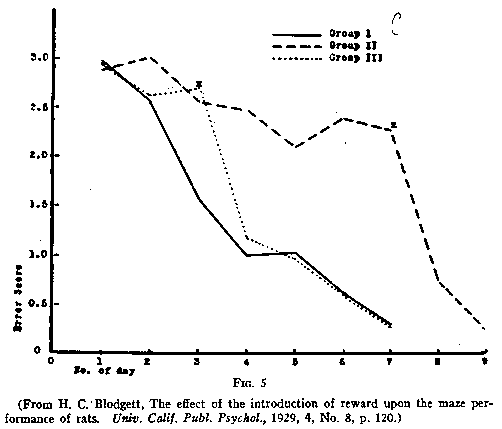
The animals of Group II, the dash line, were not fed in the maze for the first
six days but only in their home cages some two hours later. On the seventh day
(indicated by the small cross) the rats found food at the end of the maze for
the first time and continued to find it on subsequent days. The animals of Group
III were treated similarly except that they first found food at the end of the
maze on the third day and continued to find it there on subsequent days. It will
be observed that the experimental groups as long as they were not finding food
did not appear to learn much. (Their error curves did not drop.) But on the days
immediately succeeding their first finding of the food their error curves did
drop astoundingly. It appeared, in short, that during the non-rewarded trials
these animals had been learning much more than they had exhibited. This
learning, which did not manifest itself until after the food had been
introduced, Blodgett called "latent learning." Interpreting these
results anthropomorphically, we would say that as long as the animals were not
getting any food at the end of the maze they continued to take their [p.195]
time in going through it-they continued to enter many blinds. Once, however,
they knew they were to get food, they demonstrated that during these preceding
non-rewarded trials they had learned where many of the blinds were. They had
been building up a 'map,' and could utilize the latter as soon as they were
motivated to do so.
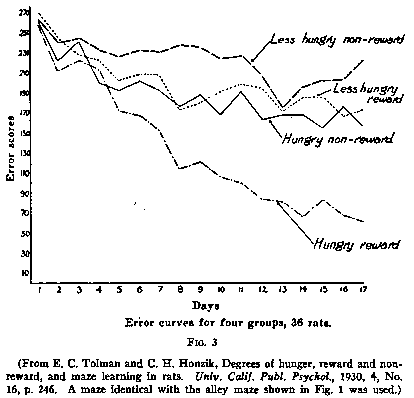
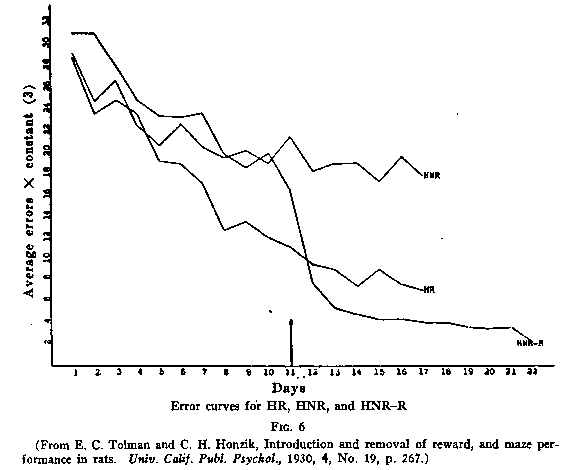
Honzik and myself repeated the experiments (or rather he did and I got some
of the credit) with the 14-unit T-mazes shown in Fig.1, and with larger groups
of animals, and got similar results. The resulting curves are shown in Fig.6. We
used two control groups-one that never found food in the maze (HNR) and one that
found it throughout (HR). The experimental group (HNR-R) found food at the end
of the maze from the 11th day on and showed the same sort of a sudden drop. But
probably the best experiment demonstrating latent learning was, unfortunately,
done not in Berkeley but at the University of Iowa, by Spence and Lippitt. Only
an abstract of this experiment has as yet been published. However, Spence has
sent a preliminary manuscript from which the following account is summarized.
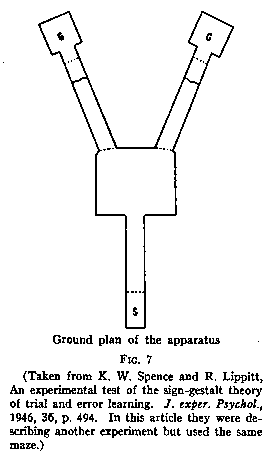
A simple Y-maze (see Fig.7) with two goal-boxes was used. Water was at the end
of the right arm of the Y and food at the end of the left arm. During the
training period the rats were run neither hungry nor thirsty. They were satiated
for both food and water before each day's trials. However, they were willing to
run because after each run they were taken out of whichever end box they had got
to and put into a living cage, with other animals in it. They were given four
trials a day in this fashion [p.196] for seven days, two trials to the right and
two to the left.
In the crucial test the animals were divided into two subgroups one made solely
hungry and one solely thirsty. It was then found that on the first trial the
hungry group went at once to the left, where the food had been, statistically
more frequently than to the right; and the thirsty group went to the right,
where the water had been, statistically more frequently than to the left. These
results indicated that under the previous non-differential and very mild
rewarding conditions of merely being returned to the home cages the animals had
nevertheless been learning where the water was and where the food was. In short,
they had acquired a cognitive map to the effect that food was to the left and
water to the right, although during the acquisition of this map they had not
exhibited any stimulus-response propensities to go more to the side which became
later the side of the appropriate goal.
There have been numerous other latent learning experiments done in the Berkeley
laboratory and elsewhere. In general, they have for the most part all confirmed
the above sort of findings.
Let us turn now to the second group of experiments.
(2) "Vicarious Trial and Error" or "VTE." The term
Vicarious Trial and Error (abbreviated as VTE) was invented by Prof. Muenzinger
at Colorado[2]
to designate the hesitating, look-[p.197] king-back-and-forth, sort of behavior
which rats can often be observed to indulge in at a choice-point before actually
going one way or the other.
Quite a number of experiments upon VTEing have been carried out in our
laboratory. I shall report only a few. In most of them what is called a
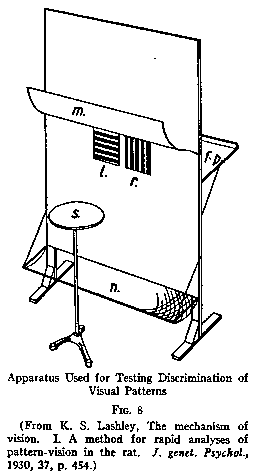
discrimination set-up has been used. In one characteristic type of visual
discrimination apparatus designed by Lashly (shown in Fig.8) the animal is put
on a jumping stand and faced with two doors which differ in some visual property
say, as here shown, vertical stripes vs. horizontal stripes.
One of each such pair of visual stimuli is made always correct and the other
wrong; and the two are interchanged from side to side in random fashion. The
animal is required to learn, say, that the vertically striped door is always the
correct one. If he jumps to it, the door falls open and he gets to food on a
platform behind. If, on the other hand, he jumps incorrectly, he finds the door
locked and falls into a net some two feet below from which he is picked up and
started over again.
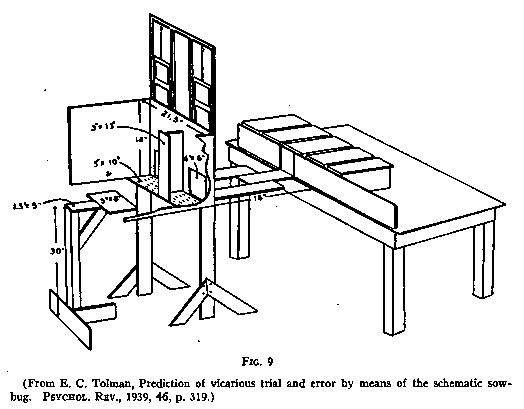
Using a similar set-up (see Fig. 9), but with landing platforms in front of the
doors so that if the rat chose incorrectly he could jump back again and start
over, I found that when the choice was an easy one, say between a white door and
a black door, the animals not only learned sooner but also did more VTEing than
when the choice was difficult, say between a white door and a gray door (see
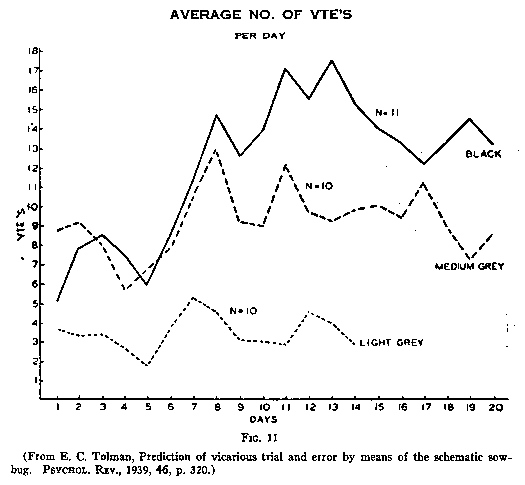
Fig.10). It appeared further (see Fig.11) that the VTEing began to appear just
as (or just before) the rats began to learn. After the learning had become
established, however, the VTE's began to go down. Further, in a study of
individual dif-[p.198] ferences by myself, Geier and Levin[3]
(actually done by Geier and Levin) using this same visual discrimination
apparatus, it was found that with one and the same difficulty of problem the
smarter animal did the more VTEing.
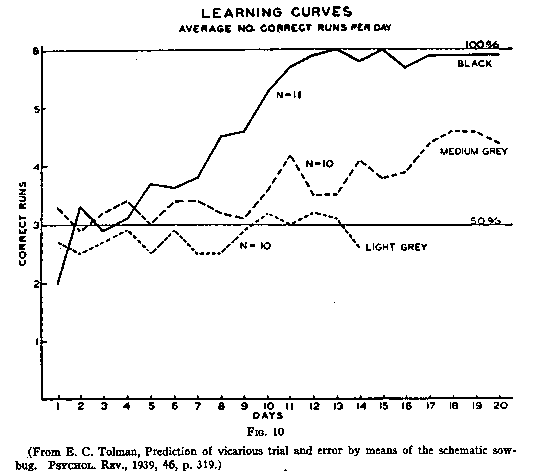
To sum up, in visual discrimination experiments the better the learning,
the more the VTE's. But this seems contrary to what we would perhaps have
expected. We ourselves would expect to do more VTEing, more sampling of the two
stimuli, when it is difficult to choose between them than when it is easy.
What is the explanation? The answer lies, I believe, in the fact that the manner
in which we set the visual discrimination problems for the rats and the manner
in which we set similar problems for ourselves are different. We already
have our 'instructions.' We know beforehand what it is we are to do. We are
told, or we tell ourselves, that it is the lighter of the two grays, the heavier
of the two weights, or the like, which is to be chosen. In such a setting we do
more sampling, more VTEing, when the stimulus-difference is small. But for the
rats the usual problem in a discrimination apparatus is quite different. They do
not know what is wanted of them. The major part of their learning in most such
experiments seems to consist in their dis-[p.199] covering the instructions. The
rats have to discover that it is the differences in visual brightness, not the
differences between left and right, which they are to pay attention to. Their
VTEing appears when they begin to 'catch on.' The greater the difference between
the two stimuli the more the animals are attracted by this difference. Hence the
sooner they catch on, and during this catching on, the more they VTE.
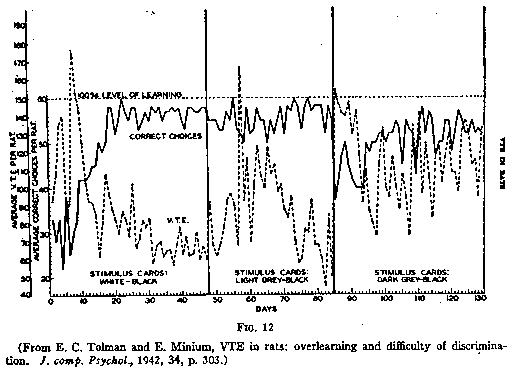
That this is a reasonable interpretation appeared further, from an experiment by
myself and Minium (the actual work done, of course, by Minium) in which a group
of six rats was first taught a white vs. black discrimination, then two
successively more difficult gray vs. black discriminations. For each difficulty
the rats were given a long series of further trials beyond the points at which
they had learned. Comparing the beginning of each of these three difficulties
the results were that the rats did more VTEing for the easy discriminations than
for the more difficult ones. When, however, it came to a comparison of amounts
of VTEing during the final performance after each learning had reached a
plateau, the opposite results were obtained. In other words, after the rats had
finally divined their instructions, then they, like human beings, did more
VTEing, more sampling, the more difficult the discrimination.
Finally, now let us note that is was [p.200] also found at Berkeley by
Jackson[4]
that in a maze the difficult maze units produce more VTEing and also that the
more stupid rats do the more VTEing. The explanation, as I see it, is that, in
the case of mazes, rats know their instructions. For them it is natural to
expect that the same spatial path will always lead to the same outcome. Rats in
mazes don't have to be told.
But what, now, is the final significance of all this VTEing? How do these
facts about VTEing affect our theoretical argument? My answer is that these
facts lend further support to the doctrine of a building up of maps. VTEing, as
I see it, is evidence that in the critical stages-whether in the first picking
up of the instructions or in the later making sure of which stimulus is
which-the animal's activity is not just one of responding passively to discrete
stimuli, but rather one of the active selecting and comparing of stimuli. The
brings me then to the third type of experiment.
(3) "Searching for the Stimulus." I refer to a recent,
and it seems to me extremely important experiment, done for a Ph.D. dissertation
by Hudson. Hudson was first interested in the question of whether or not rats
could learn an avoidance reaction in one trial. His animals were tested one at a
time in a living cage (see Fig.13) with a small striped visual pattern at the
end, on which was mounted a food cup. The hungry rat approached this food cup
and ate. An electrical arrangement was provided so that when the rat touched the
cup he could be given an electric shock. And one such shock did appear to be
enough. For when the rat was replaced in this same cage days or even weeks
afterwards, he usually demonstrated immediately strong avoidance reactions to
the visual pattern. The animal withdrew from that end of the [p.201] cage, or
piled up sawdust and covered the pattern, or showed various other amusing
responses all of which were in the nature of withdrawing from the pattern or
making it disappear.
But the particular finding which I am interested in now appeared as a result
of a modification of this standard procedure. Hudson noticed that the animals,
anthropomorphically speaking, often seemed to look around after the shock
to see what it was that had hit them. Hence it occurred to him that, if the
pattern were made to disappear the instant the shock occurred, the rats might
not establish the association. And this indeed is what happened in the case of
many individuals. Hudson added further electrical connections so that when the
shock was received during the eating, the lights went out, the pattern and the
food cup dropped out of sight, and the lights came on again all within the
matter of a second. When such animals were again put in the cage 24 hours later,
a large percentage showed no avoidance of the pattern. Or to quote Hudson's own
words:
"Learning what object to avoid...may occur exclusively during the period
after the shock. For if the object from which the shock was actually
received is removed at the moment of the shock, a significant number of animals
fail to learn to avoid it, some selecting other features in the environment for
avoidance, and others avoiding nothing."

In other words, I feel that this experiment reinforces the notion of the largely
active selective character in the rat's building up of his cognitive map. He
often has to look actively for the significant stimuli in order to form his map
and does not merely passively receive and react to all the stimuli which are
physically present.
Turn now to the fourth type of experiment.
(4) The "Hypothesis" Experiments. Both the notion of hypotheses
in rats and the design of the experiments to demonstrate such hypotheses are to
be credited to Krech. Krech used a four-compartment discrimination-box. In such
a four-choice box the correct door at each choice-point may be determined by the
experimenter in terms of its being lighted or dark, left or right, or various
combinations of these. If all [p.202] possibilities are randomized for the 40
choices made in 10 runs of each day's test, the problem could be made insoluble.
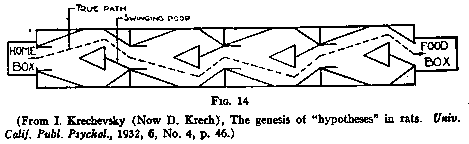
When this was done, Krech found that the individual rat went through a
succession of systematic choices. That is, the individual animal might perhaps
begin by choosing practically all right-handed doors, then he might give this up
for choosing practically all left-hand doors, and then, for choosing all dark
doors, and so on. These relatively persistent, and well-above-chance systematic
types of choice Krech called "hypotheses." In using this term he
obviously did not mean to imply verbal processes in the rat but merely referred
to what I have been calling cognitive maps which, it appears from his
experiments, get set up in a tentative fashion to be tried out first one and
then another until, if possible, one is found which works.
Finally, it is to be noted that these hypothesis experiments, like the latent
learning, VTE, and "looking for the stimulus" experiments, do not, as
such, throw light upon the widths of the maps which are picked up but do
indicate [p.203] the generally map-like and self-initiated character of
learning.
For the beginning of an attack upon the problem of the width of the maps let
me turn to the last group of experiments.
(5) "Spatial Orientation" Experiments. As early as 1929,
Lashley reported incidentally the case of a couple of his rats who, after having
learned an alley maze, pushed back the cover near the starting box, climbed out
and ran directly across the top to the goal-box where they climbed down in again
and ate. Other investigators have reported related findings. All such
observations suggest that rats really develop wider spatial maps which include
more than the mere trained-on specific paths. In the experiments now to be
reported this possibility has been subjected to further examination.
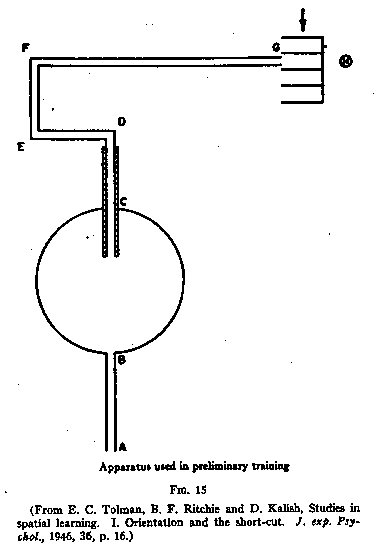
In the first experiment, Tolman, Ritchie and Kalish (actually Ritchie and
Kalish) used the set-up shown in Fig.15.
This was an elevated maze. The animals ran from A across the open circular table
through CD (which had alley walls) and finally to G, the food box. H was a light
which shone directly down the path from G to F. After four nights, three trials
per night, in which the rats learned to run directly and without hesitation from
A to G, the apparatus was changed to the sun-burst shown in Fig.16. The starting
path and the table remained the same but a series of radiating paths was added.
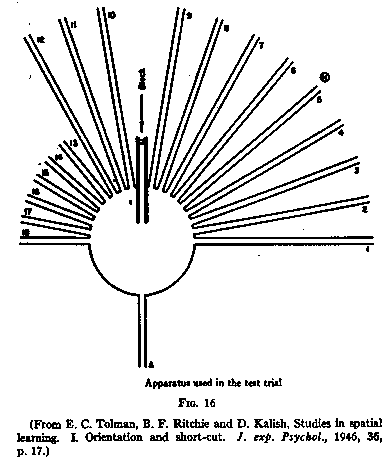
The animals were again started at A [p.204] and ran across the circular table
into the alley and found themselves blocked. They then returned onto the table
and began exploring practically all the radiating paths. After going out a few
inches only on any one path, each rat finally chose to run all the way out on
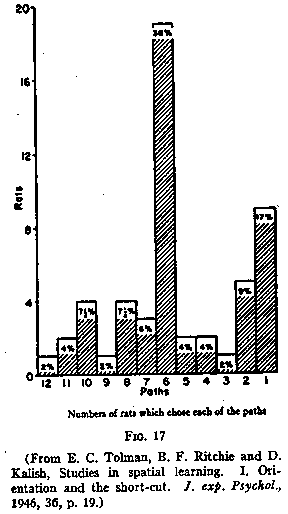
one. The percentages of rats finally choosing each of the long paths from 1 to
12 are shown in Fig.17. It appears that there was a preponderant tendency to
choose path No.6 which ran to a point some four inches in front of where the
entrance to the food-box had been. The only other path chosen with any
appreciable frequency was No.1-that is, the path which pointed perpendicularly
to the food-side of the room.
These results seem to indicate that the rats in this experiment had learned
not only to run rapidly down the original roundabout route but also, when this
was blocked and radiating paths presented, to select one pointing rather
directly towards the point where the food had been or else at least to select a
path running perpendicularly to the food-side of the room.
As a result of their original training, the rats had, it would seem, acquired
not merely a strip-map to the effect that the original specifically trained-on
path led to food but, rather, a wider comprehensive map to the effect that food
was located in such and such a direction in the room.
Consider now a further experiment done by Ritchie alone. This experiment
tested still further the breadth of the spatial map which is acquired. In this
further experiment the rats were again run across the table-this time to the
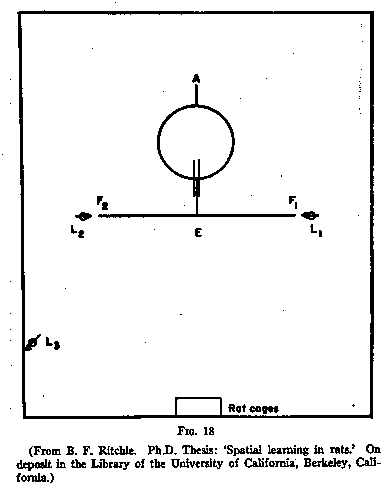
arms of a simple T. (See Fig.18.)
Twenty-five animals were trained for seven days, 20 trials in all, to find
food at F1; and twenty-five animals were trained to find it at F2.
The L's in the diagram indicate lights. On the eighth day the starting path and
table top were rotated through 180 degrees so that they were now in the position
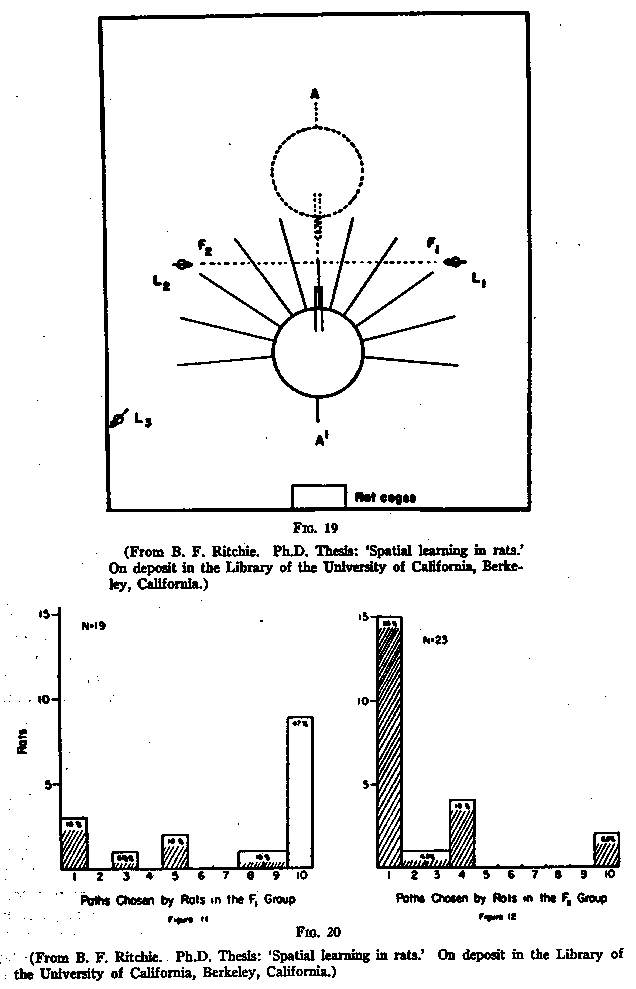
shown in Fig.19. The dottted lines represent the old position. And a series of
radiating paths was added. What happened? Again the rats ran across the table
into the central alley. When, however, they found themselves blocked, they
turned back onto the table and this time also spent many seconds touching and
trying out for only a few steps practically all the paths. Finally, however,
within seven minutes, 42 of the 50 rats chose one path and ran all the way out
on it. The paths finally chosen by the 19 of these animals that had been fed at
F1 and by the 23 that had been fed at F2 are shown in
Fig.20. [p.205]
This time the rats tended to choose, not the paths which pointed directly to the
spots where the food had been, but rather paths which ran perpendicularly to the
corresponding sides of the room. The spatial maps of these rats, when the
animals were started from the opposite side of the room, were thus not
completely adequate to the precise goal positions but were adequate as to the
correct sides of the room. The maps of these animals were, in short, not
altogether strip-like and narrow.
This completes my report of experiments. There were the latent learning
experiments, the VTE experiments, the searching for the stimulus experiment, the
hypothesis experiments, and these last spatial orientation experiments.
And now, at last, I come to the humanly significant and exciting problem:
namely, what are the conditions which favor narrow strip-maps and what are those
which tend to favor broad comprehensive maps not only in rats but also in men?
There is considerable evidence scattered throughout the literature bearing on
this question both for rats and for men. Some of this evidence was obtained in
Berkeley and some of it elsewhere. I have not time to present it in any detail.
I can merely summarize it by saying that narrow strip maps rather than broad
comprehensive maps seem to be induced: (1) by a damaged brain, (2) by an
inadequate array of environmentally presented cues, (3) by an [p.206]
[p.207] overdose of repetitions on the original trained-on path and (4) by the
presence of too strongly motivational or of too strongly frustrating conditions.
It is this fourth factor which I wish to elaborate upon briefly in my
concluding remarks. For it is going to be my contention that some, at least, of
the so-called 'psychological mechanisms' which the clinical psychologists and
the other students of personality have uncovered as the devils underlying many
of our individual and social malajustments can be interpreted as narrowings of
our cognitive maps due to too strong motivations or to too intense frustration.
My argument will be brief, cavalier, and dogmatic. For I am not myself a
clinician or a social psychologist. What I am going to say must be considered,
therefore, simply as in the nature of a rat psychologist's ratiocinations
offered free.
By way of illustration, let me suggest that at least the three dynamisms
called, respectively, "regression," "fixation," and
"displacement of aggression onto outgroups" are expressions of
cognitive maps which are too narrow and which get built up in us as a result of
too violent motivation or of too intense frustration.
(a) Consider regression. This is the term used for those cases in
which an individual, in the face of too difficult a problem, returns to earlier
more childish ways of behaving. Thus, to take an example, the overprotected
middle-aged woman (reported a couple of years ago in Time Magazine) who,
after losing her husband, regressed (much to the distress of her growing
daughters) into dressing in too youthful a fashion and into competing for their
beaux and then finally into behaving like a child requiring continuous care,
would be an illustration of regression. I would not wish you to put too much
confidence in the reportorial accuracy of Time, but such an extreme case
is not too different from many actually to be found in our mental hospitals or
even sometimes in ourselves. In all such instances my argument would be (1) that
such regression results from too strong a present emotional situation and (2)
that it consists in going back to too narrow an earlier map, itself due to too
much frustration or motivation in early childhood. Time's middle-aged
woman was presented by too frustrating an emotional situation at her husband's
death and she regressed, I would wager, to too narrow adolescent and childhood
maps since these latter had been originally excessively impressed because of
overstressful experiences at the time she was growing up.
(b) Consider fixation. Regression and fixation tend to go hand in
hand. For another way of stating the fact of the undue persistence of early maps
is to say that they were fixated. This has even been demonstrated in rats. If
rats are too strongly motivated in their original learning, they find it very
difficult to relearn when the original path is no longer correct. Also after
they have relearned, if they are given an electric shock they, like Time's
woman, tend to regress back again to choosing the earlier path.
(c) Finally, consider the "displacement of aggression onto
outgroups." Adherence to one's own group is an ever-present tendency
among primates. It is found in chimpanzees and monkeys as strongly as in men. We
primates operate in groups. And each individual in such a group tends to
identify with his whole group in the sense that the group's goal's become his
goals, the group's life and immortality, his life and immortality. Furthermore,
each individual soon learns that, when as an individual he is frustrated, he
must not take out his aggressions on [p.208] the other members of his own group.
He learns instead to displace his aggressions onto outgroups. Such a
displacement of aggression I would claim is also a narrowing of the cognitive
map. The individual comes no longer to distinguish the true locus of the cause
of his frustration. The poor Southern whites, who take it out on the Negroes,
are displacing their aggressions from the landlords, the southern economic
system, the northern capitalists, or wherever the true cause of their
frustration may lie, onto a mere convenient outgroup. The physicists on the
Faculty who criticize the humanities, or we psychologists who criticize all the
other departments, or the University as a whole which criticizes the Secondary
School system or, vice versa, the Secondary School system which criticizes the
University-or, on a still larger and far more dangerous scene-we Americans who
criticize the Russians and the Russians who criticize us, are also engaging, at
least in part, in nothing more than such irrational displacements of our
aggressions onto outgroups.
I do not mean to imply that there may not be some true interferences by the
one group with the goals of the other and hence that the aggressions of the
members of the one group against the members of the other are necessarily wholly
and merely displaced aggressions. But I do assert that often and in
large part they are such mere displacements.
Over and over again men are blinded by too violent motivations and too
intense frustrations into blind and unintelligent and in the end desperately
dangerous hates of outsiders. And the expression of these their displaced hates
ranges all the way from discrimination against minorities to world
conflagrations.
What is the name of Heaven and Psychology can we do about it? My only answer
is to preach again the virtues of reason-of, that is, broad cognitive maps. And
to suggest that the child-trainers and the world-planners of the future can
only, if at all, bring about the presence of the required rationality (i.e.,
comprehensive maps) if they see to it that nobody's children are too
over-motivated or too frustrated. Only then can these children learn to look
before and after, learn to see that there are often round-about and safer paths
to their quite proper goals-learn, that is, to realize that the well-beings of
White and of Negro, of Catholic and of Protestant, of Christian and of Jew, of
American and of Russian (and even of males and females) are mutually
interdependent.
We dare not let ourselves or others become so over-emotional, so hungry, so
ill-clad, so over-motivated that only narrow strip-maps will be developed. All
of us in Europe as well as in America, in the Orient as well as in the Occident,
must be made calm enough and well-fed enough to be able to develop truly
comprehensive maps, or, as Freud would have put it, to be able to learn to live
according to the Reality Principle rather than according to the too narrow and
too immediate Pleasure Principle.
We must, in short, subject our children and ourselves (as the kindly
experimenter would his rats) to the optimal conditions of moderate motivation
and of an absence of unnecessary frustrations, whenever we put them and
ourselves before that great God-given maze which is our human world. I cannot
predict whether or not we will be able, or be allowed, to do this; but I can say
that, only insofar as we are able and are allowed, have we cause
for hope.
Footnotes
[1] 34th Annual Faculty Research Lecture, delivered at the
University of California, Berkeley, March 17, 1947. Presented also on March 26,
1947 as one in a series of lectures in Dynamic Psychology sponsored by the
division of psychology of Western Reserve University, Cleveland, Ohio.
[2] Vide: K.F. Muenzinger, Vicarious trial and error at a
point of choice: I.A general survey of its relation to learning efficiency. J.genet.
Psychol., 1938, 53, 75-86.
[3] F.M. Geier, M. Levin & E.C. Tolman, Individual
differences in emotionality, hypothesis formation, vicarious trial and error and
visual discrimination learning in rats. Comp. Psychol. Monor., 1941, 17,
No.3.
[4] L.L. Jackson, V.T.E. on an elevated maze. J.comp.
Psychol., 1943, 36, 99-107.